先史時代の両生類の写真とプロフィール

Nobu Tamura
石炭紀とペルム紀の間、 先史時代の両生類 、爬虫類ではなく、地球の大陸の頂点捕食者でした。次のスライドでは、両生類からウエストロチアナまで、30 を超える先史時代の両生類の写真と詳細なプロフィールをご覧いただけます。
01/33両生類

アラン・ベネトー
生物のファミリーにその名前を付ける属が、そのファミリーの中で最も理解されていないメンバーであることがよくあります。 Amphibamus の場合、話はもう少し複雑です。 「両生類」という言葉は、有名な古生物学者が エドワード・ドリンカー・コープ 後期の化石にこの名前を付けた石炭紀限目。両生類は、この時点で陸生生物を支配していた大型のクロコダイルのような「テムノスポンディル」両生類 (エリオプスやマストドンサウルスなど) のはるかに小さいバージョンであったようですが、カエルやサンショウウオが進化の歴史のポイントを表している可能性もあります。両生類の系統樹から分かれます。いずれにせよ、アンピバムスは小型で無害な生き物であり、最近の四足動物の祖先よりもわずかに洗練されていました.
02/33アルケゴサウルス
 Nobu Tamura
Nobu Tamura ' id='mntl-sc-block-image_2-0-5' />
Nobu Tamura
アルケゴサウルスの完全な頭骨と部分的な頭骨がどれだけ多く発見されたかを考えると、ほぼ 200 頭で、そのすべてがドイツの同じ化石サイトから出土していますが、これはまだ比較的神秘的な先史時代の両生類です。再構成から判断すると、アルケゴサウルスは西ヨーロッパの沼地をうろつき、小魚や (おそらく) 小型の両生類や テトラポッド .ちなみに、「アーキゴサウルス科」の傘の下には、さらにあいまいな両生類がいくつかあり、そのうちの 1 つはコリドスクスという面白い名前が付けられています。
03/33ベルゼブフォ(デビルフロッグ)
 米国科学アカデミー
米国科学アカデミー ' id='mntl-sc-block-image_2-0-9' />
米国科学アカデミー
の 白亜紀ベルゼブフォ 体重約 10 ポンド、頭から尾まで 1 フィート半の、これまでに生きた中で最大のカエルでした。異常に広い口で、通常の大きな昆虫の食事だけでなく、時折恐竜の赤ちゃんを食べていたのでしょう。
04/33ブランキオサウルス

Nobu Tamura
たった一文字でこんなに違うなんて驚きです。 ブラキオサウルス これまで地球を歩き回った最大の恐竜の 1 つでしたが、ブランキオサウルス (1 億 5000 万年前に生きていた) は、すべての先史時代の両生類の中で最小の 1 つでした。この6インチの長さの生き物は、以前はより大きな「テムノスポンディル」両生類(エリオプスなど)の幼虫期を代表していると考えられていましたが、それ自体の属に値すると考える古生物学者の数が増えています.いずれにせよ、ブランキオサウルスは、より大きなテモンスポンディルのいとこの解剖学的特徴をミニチュアで備えており、最も顕著なのは特大のほぼ三角形の頭です。
05/33カコップ
 フィールド自然史博物館
フィールド自然史博物館 ' id='mntl-sc-block-image_2-0-16' />
フィールド自然史博物館
最も初期の両生類の中でより爬虫類に似たカコップスは、ずんぐりした足、短い尾、軽装甲の背中を持つ、ずんぐりした猫サイズの生き物でした。この先史時代の両生類は比較的高度な鼓膜 (陸上での生活に必要な適応) を持っていたという証拠がいくつかあります。 二畳紀 北アメリカの生息地 (太陽の灼熱の熱だけでなく)。
06/33コロステウス
 Nobu Tamura' id='mntl-sc-block-image_2-0-20' />
Nobu Tamura' id='mntl-sc-block-image_2-0-20' /> Nobu Tamura
何億年も前の石炭紀の時代、葉鰭が発達した進化した魚類、最初の陸上冒険型四足動物、そして最も原始的な両生類を区別することは非常に困難でした。オハイオ州に多くの化石が見られるコロステウスは、しばしば四足動物として説明されますが、ほとんどの古生物学者は、この生き物を「コロステウス」両生類として分類することに慣れています。コロステウスの長さは約 3 フィートで、足は非常に小さく (役に立たないとは言えません)、平らで先のとがった頭にはそれほど脅威ではない 2 つの牙がありました。おそらくほとんどの時間を水中で過ごし、小さな海の動物を食べていたのでしょう。
07/33サイクロトサウルス

Nobu Tamura
両生類の黄金時代は、面白い名前のマストドンサウルスに代表される巨大な沼地に住む「テムノスポンディル」の家族によってもたらされました。マストドンサウルスの近縁種であるサイクロトサウルスの化石は、西ヨーロッパからグリーンランド、タイに至るまで、異常に広い地理的範囲で発見されており、私たちが知る限り、テムノスポンディルの最後の 1 つでした。 (両生類は、2000 年代初頭までに個体数が減少し始めました。 ジュラ紀 期間、今日も続く下降スパイラル。)
マストドンサウルスと同様に、サイクロトサウルスの最も注目すべき特徴は、その大きくて平らなワニのような頭であり、比較的ちっぽけな両生類の幹に取り付けられたとき、漠然と気まぐれに見えました。当時の他の両生類と同様に、サイクロトサウルスはおそらく海岸線をうろつき、さまざまな海洋生物 (魚、軟体動物など) や、時折小さなトカゲや哺乳類を捕まえることで生計を立てていました。
08/33ディプロカウルス
 ウィキメディア・コモンズ' id='mntl-sc-block-image_2-0-29' />
ウィキメディア・コモンズ' id='mntl-sc-block-image_2-0-29' /> ウィキメディア・コモンズ
ディプロカウルスはそれらの古代の 1 つです。 両生類 箱から出してすぐに間違って組み立てられたように見えます。比較的平らで目立たない幹が、両側にブーメラン型の骨の突起で飾られた非常に特大の頭に取り付けられています。ディプロカウルスの頭骨はなぜこれほど珍しいものだったのですか?考えられる説明は 2 つあります。その V 字型のノギンは、この両生類が強い海流や川の流れをナビゲートするのに役立った可能性があります。また、その巨大な頭が、後期の大型の海洋捕食者の食欲をそそった可能性もあります。 二畳紀 これにより、獲物をより簡単に飲み込むことができなくなりました。
09/33エオカエシリア

Nobu Tamura
両生類の主要な 3 科の名前を尋ねられたとき、ほとんどの人はカエルとサンショウウオを簡単に思いつくでしょうが、アシナシイモリを思い浮かべる人は多くありません。 Eocaecilia は、化石記録でまだ確認されている最古のアシナシイモリです。実際、この属は非常に「基本的」であったため、小さな痕跡の足をまだ保持していました (最も初期の 先史時代のヘビ 白亜紀の)。どちら(完全な足)について 先史時代の両生類 エオカエキリアから進化したのかは謎のまま。
10/33エオギリヌス

Nobu Tamura
眼鏡をかけずにエオギリノスを見た場合、これを誤解した可能性があります 先史時代の両生類 良いサイズのヘビの場合。ヘビのように鱗で覆われており(魚の祖先から直接受け継いだもの)、後期の沼地をねじれて進む際に保護するのに役立ちました。石炭紀限目。エオギリヌスには短く太い脚があり、この初期の両生類は半水生のクロコダイルのようなライフスタイルを追求し、浅瀬から小さな魚を捕まえていたようです.
11/33エリオプス

ウィキメディア・コモンズ
最もよく知られているの 1 つ 先史時代の両生類 初期の 二畳紀 期間、エリオプスは大まかな輪郭を持っていました クロコダイル 、その低めの胴体、広げられた足、そして巨大な頭を持っています。当時最大の陸生動物の 1 つであったエリオプスは、その後に続いた真の爬虫類と比べてそれほど大きくはなく、体長はわずか 6 フィート、200 ポンドにすぎませんでした。おそらくワニのように狩りをし、浅い湿地の水面のすぐ下に浮かんで、近づきすぎた魚を捕まえました。
12/33フェデシア
 カーネギー自然史博物館' id='mntl-sc-block-image_2-0-45' />
カーネギー自然史博物館' id='mntl-sc-block-image_2-0-45' /> カーネギー自然史博物館
Fedexia は、一部の企業スポンサー プログラムのルーブリックの下では名前が付けられていませんでした。むしろ、この 3 億年前の両生類の化石は、ピッツバーグ国際空港のフェデラル エクスプレス グラウンド本部の近くで発見されました。しかし、その特徴的な名前を除けば、Fedexia はプレーン バニラ タイプのようです。 先史時代の両生類 、生い茂ったサンショウウオを漠然と連想させ、(歯の大きさと形から判断すると)後期の小さな虫や陸上動物を食べて生きています石炭紀限目。
13/33胃で陰気なカエル

ウィキメディア・コモンズ
その名前が示すように、胃陰気なカエルは、幼生を妊娠するための奇妙な方法を持っていました. メスは、オタマジャクシが食道を介して登る前に、胃の中で安全に発達した新しく受精した卵を飲み込みました.見る 胃陰気なカエルの詳細なプロファイル
14/33ゲロバトラクス
 ウィキメディア・コモンズ' id='mntl-sc-block-image_2-0-52' />
ウィキメディア・コモンズ' id='mntl-sc-block-image_2-0-52' /> ウィキメディア・コモンズ
2 億 9000 万年前の生物の 1 つの不完全な化石が、いかに古生物学の世界を揺るがすことができるかは驚くべきことです。 2008 年にデビューしたとき、ゲロバトラクスは「フロガマンダー」として広く宣伝されました。これは、カエルとサンショウウオの最後の共通の祖先であり、現代の両生類の 2 つの最も人口の多い家族です。 (公平を期すために言うと、ゲロバトラコスの大きなカエルのような頭骨と、その比較的細いサンショウウオのような体との組み合わせは、どんな科学者も考えさせます。)これが意味することは、カエルとサンショウウオが数百万年後に別々の道を歩んだということです両生類の進化の既知の速度を大幅に加速するゲロバトラクスの時代。
15/33ゲロソラックス
 ウィキメディア・コモンズ' id='mntl-sc-block-image_2-0-56' />
ウィキメディア・コモンズ' id='mntl-sc-block-image_2-0-56' /> ウィキメディア・コモンズ
すべての先史時代の両生類の中で最も特徴的なものの 1 つである Gerrothorax は、上に目が固定された平らなフットボール型の頭と、首から突き出た外側の羽のような鰓を持っていました。これらの適応は、ジェロソラックスが(すべてではないにしても)ほとんどの時間を水中で過ごしたこと、そしてこの両生類が独特の狩猟戦略を持っていた可能性があるという確かな手がかりです。口。おそらく、他の海の捕食者に対する保護の形として、後期 三畳紀 ジェロソラックスはまた、体の上部と下部に沿って軽装甲の皮膚を持っていました.
16/33黄金のヒキガエル

米国魚類野生生物局
野生で最後に確認されたのは 1989 年で、コスタリカの他の場所で奇跡的に個体が発見されない限り、絶滅したと推定されています。 黄金のヒキガエル 両生類の個体数が世界的に減少している謎の原因となっています。
17/33Karaurus
 ウィキメディア・コモンズ' id='mntl-sc-block-image_2-0-63' />
ウィキメディア・コモンズ' id='mntl-sc-block-image_2-0-63' /> ウィキメディア・コモンズ
古生物学者によって、最初の真のサンショウウオ (または少なくとも、化石が発見された最初の真のサンショウウオ) であると考えられているカラウルスは、両生類の進化の比較的後期に出現し、 ジュラ紀 限目。将来の化石の発見によって、ペルム紀と三畳紀のより大きく恐ろしい祖先からのこの小さな生物の発達に関するギャップが埋められる可能性があります。
18/33クーラスクス

ウィキメディア・コモンズ
クーラスクスの最も注目すべき点は、このオーストラリアの両生類が生きていた時期です。白亜紀中期、またはマストドンサウルスのような有名な「テムノスポンディル」の祖先が北半球で絶滅してから約 1 億年後です。クーラスクスは、クロコダイルのような基本的な脊椎の体型(特大の頭としゃがんだ手足のある長い胴体)を守っており、魚と甲殻類の両方を食べていたようです。クーラスクスは、北方の近縁種が地上から姿を消した後、どのようにして繁栄したのでしょうか?おそらく、白亜紀のオーストラリアの涼しい気候がそれと関係があり、クーラスクスが長期間冬眠し、捕食を避けることができた.
19/33マストドンサウルス

ドミトリー・ボグダノフ
確かに、「マストドンサウルス」はクールな響きの名前ですが、「マストドン」がギリシャ語で「乳歯」を意味することを知っていれば、あまり印象に残らないかもしれません (もちろん、氷河期にも当てはまります)。 マストドン 同じように)。マストドンサウルスは、これまでに生きた中で最大の先史時代の両生類の1つであり、体全体のほぼ半分の長さである巨大で細長い平らな頭を持つ奇妙なプロポーションの生き物でした.大きくて不格好な胴体とずんぐりした脚を考えると、三畳紀後期のマストドンサウルスがすべての時間を水中で過ごしたのか、それともおいしいおやつを求めてときどき乾いた土地に足を踏み入れたのかは不明です。
20/33メガロケファルス

ドミトリー・ボグダノフ
その名前(「巨大な頭」のギリシャ語)が印象的であるのと同じくらい印象的ですが、メガロケファルスは石炭紀後期の比較的あいまいな先史時代の両生類のままです。それについて私たちが知っていることのほとんどは、それが巨大な頭を持っていたということだけです.それでも、古生物学者は、メガロケファルスがワニのような体格を持っていたと推測できます。 先史時代のワニ 同様に、そのずんぐりした足で湖岸や川床をうろつき、近くをさまよっている小さな生き物を捕まえます。
21/33メトポサウルス
 ウィキメディア・コモンズ' id='mntl-sc-block-image_2-0-79' />
ウィキメディア・コモンズ' id='mntl-sc-block-image_2-0-79' /> ウィキメディア・コモンズ
石炭紀とペルム紀の長い間、巨大な両生類は地球上で支配的な陸上動物でしたが、その長い統治は2億年前の三畳紀の終わりまでに終わりました.この品種の典型的な例は、奇妙な特大の平らな頭と長い魚のような尾を持つワニのような捕食者であるメトポサウルスでした。四足歩行の姿勢 (少なくとも陸上では) と比較的弱い手足を考えると、メトポサウルスは人類に大きな脅威を与えることはなかったでしょう。 初期の恐竜 北アメリカと西ヨーロッパ (そしておそらく世界の他の地域も同様) の浅い湿地と湖で魚を食べながら共存していました。
その奇妙な解剖学から、メトポサウルスは明らかに特殊なライフスタイルを追求したにちがいない。一説によると、この 0.5 トンの両生類は、浅い湖の水面近くを泳ぎ、これらの水域が乾くと、湿った土に潜り込み、雨季が戻るまでの時間を待っていました。 (この仮説の問題点は、三畳紀後期の他のほとんどの穴を掘る動物がメトポサウルスのサイズのほんの一部だったことです。) それが大きかったとしても、メトポサウルスは捕食の影響を受けませんでした。フィトサウルスの家族 ワニのような爬虫類 それはまた、半水生の存在を導きました。
22/33マイクロブラチス

Nobu Tamura
マイクロブラチスは、「マイクロサウルス」として知られる先史時代の両生類の中で最も注目すべき属であり、ご想像のとおり、その小さなサイズが特徴です。両生類の場合、Microbrachis はその多くの特徴を保持しています。 魚 ほっそりしたウナギのような体とちっぽけな手足など、テトラポッドの祖先。その解剖学的構造から判断すると、Microbrachis はペルム紀初期にヨーロッパの広い地域を覆う湿地で、すべてではないにしてもほとんどの時間を過ごしたようです。
23/33オフィデルペトン
 アラン・ベネトー' id='mntl-sc-block-image_2-0-88' />
アラン・ベネトー' id='mntl-sc-block-image_2-0-88' /> アラン・ベネトー
ヘビが数千万年後に進化したことを知らなかったとしたら、オフィデルペトンをシューという音を立ててとぐろを巻く生き物の 1 つと間違えがちです。真の爬虫類ではなく先史時代の両生類である Ophiderpeton とその 'astopod' の親戚は、非常に早い時期 (約 3 億 6000 万年前) に仲間の両生類から分岐したようであり、生きている子孫を残しませんでした。この属は、その細長い背骨 (200 以上の椎骨で構成されていた) と、前方を向いた目を持つ鈍い頭骨によって特徴付けられました。これは、石炭紀の生息地の小さな昆虫に適応するのに役立ちました。
24/33ペロロセファルス
 ウィキメディア・コモンズ)' id='mntl-sc-block-image_2-0-92' />
ウィキメディア・コモンズ)' id='mntl-sc-block-image_2-0-92' /> ウィキメディア・コモンズ)
ギリシア語で「巨大な頭」という意味の名前にもかかわらず、ペロロケファルスは実際にはかなり小さかったが、体長 3 フィートで、三畳紀後期の南アメリカで最大の先史時代の両生類の 1 つであった (この地域が最初の恐竜を生み出していた当時)。 )。ペロロケファルスの真の重要性は、三畳紀末の絶滅を生き延び、ジュラ紀と白亜紀に存続した数少ない両生類の1つである「チグチサウルス」であったということです。その後期の中生代の子孫は、印象的なクロコダイルのようなプロポーションに成長しました.
25/33フレゲトンティア
ウィキメディア・コモンズ
訓練されていない目には、ヘビのような先史時代の両生類であるフレゲトンティアは、同じく小さな (ぬるぬるした) ヘビに似ていたオフィデルペトンと見分けがつかないように見えるかもしれません。しかし、後期石炭紀のフレゲトンティアは、手足がないだけでなく、現代のヘビの頭蓋骨に似た異常で軽量な頭蓋骨を持っていることで、両生類の群れとは一線を画していました(この特徴は、収束進化によって説明される可能性が最も高いです)。
26/33Platyhystrix
 Nobu Tamura' id='mntl-sc-block-image_2-0-100' />
Nobu Tamura' id='mntl-sc-block-image_2-0-100' /> Nobu Tamura
ペルム紀初期の他の点では目立たない先史時代の両生類である Platyhystrix は、 ディメトロドン 背に帆のような帆があり、(他の帆船と同様に) 温度調節装置と性的に選択された特徴の 2 つの役割を果たした可能性があります。その顕著な特徴を超えて、Platyhystrix はほとんどの時間を北アメリカ南西部の湿地ではなく陸上で過ごし、昆虫や小動物を食べて生きていたようです。
27/33プリオノスクス
 ドミトリー・ボグダノフ' id='mntl-sc-block-image_2-0-104' />
ドミトリー・ボグダノフ' id='mntl-sc-block-image_2-0-104' /> ドミトリー・ボグダノフ
まず第一に、プリオノスクスが独自の属に値することに誰もが同意するわけではありません。一部の古生物学者は、この巨大な (体長約 30 フィート) 先史時代の両生類は、実際にはプラティオポサウルスの一種であると主張しています。そうは言っても、プリオノスクスは両生類の中で真の怪物であり、多くの想像上の「誰が勝つか?プリオノスクス対 [ここに大きな動物を挿入] の議論がインターネット上で行われました。もしあなたが十分に近づくことができたなら、そしてあなたが望んでいないのであれば、プリオノスクスはおそらく数千万年後に進化した大きなワニと見分けがつかず、両生類ではなく真の爬虫類だったでしょう.
28/33プロテロジリヌス
 Nobu Tamura' id='mntl-sc-block-image_2-0-108' />
Nobu Tamura' id='mntl-sc-block-image_2-0-108' /> Nobu Tamura
可能性は低いと思われるかもしれませんが、1 億年後に続いた恐竜を考えると、体長 3 フィートのプロテロギリヌスは、地球の大陸に人が住み始めたばかりの石炭紀後期ユーラシアと北アメリカの頂点捕食者でした。空気を吸う先史時代の両生類によって。プロテロギリヌスは四足動物の祖先の進化の痕跡をいくつか持っており、最も顕著なのは幅の広い魚のような尾で、その細い体の残りの部分とほぼ同じ長さでした。
29/33シーモリア
 ウィキメディア・コモンズ' id='mntl-sc-block-image_2-0-112' />
ウィキメディア・コモンズ' id='mntl-sc-block-image_2-0-112' /> ウィキメディア・コモンズ
Seymouria は明らかに水陸両用ではないように見える先史時代の両生類でした。この小さな生き物の頑丈な脚、筋肉質の背中、そして (おそらく) 乾燥した皮膚は、1940 年代の古生物学者に真の爬虫類として分類するよう促し、その後、所属する両生類のキャンプに戻った.その化石が発見されたテキサス州の町にちなんで名付けられたシーモリアは、約 2 億 8000 万年前のペルム紀初期の日和見主義的なハンターであり、昆虫、魚、その他の小さな両生類を求めて乾燥した土地や暗い沼地を歩き回っていたようです。
シームリアの肌がぬるぬるではなくうろこ状だったのはなぜですか?それが生きていた当時、北アメリカのこの地域は異常に暑く乾燥していたので、典型的な湿った皮膚の両生類は、地質学的に言えば、あっという間にしわが寄って死んでいたでしょう. (興味深いことに、セイモリアは爬虫類に似た別の特徴を持っていた可能性があります。それは、鼻の腺から余分な塩分を排出する能力です。)両生類なので、卵を産むために水に戻らなければなりませんでした。
数年前、シーモリアは BBC シリーズにカメオ出演しました。 モンスターと歩く 、おいしい食事を獲得することを期待して、ディメトロドンの卵のクラッチに潜んでいます。おそらく、この番組の R 指定のエピソードにより適しているのは、ドイツでの「タンバッハ愛好家」の発見でしょう。シーモリアの成体のペア、男性 1 人、女性 1 人が死後、並んで横たわっています。もちろん、このデュオが交尾行為の後(または交尾中に)死亡したかどうかはわかりませんが、興味深いテレビになることは間違いありません!
33 の 30ソレノドンサウルス

ドミトリー・ボグダノフ
最も進化した両生類と最も初期の真の爬虫類を区別する明確な境界線はありませんでした。さらに紛らわしいことに、これらの両生類は「より進化した」従兄弟と共存し続けました。一言で言えば、それがソレノドンサウルスを非常に混乱させるものです.たとえば、ソレノドンサウルスは非常に両生類に似た骨格を持っていましたが、その歯と内耳の構造は、水に住むいとこの特徴がありませんでした。その最も近い親戚は、はるかによく理解されているディアデクテスだったようです。
31/33トリアドバトラコス

ウィキメディア・コモンズ
より古い候補が最終的に発見される可能性はありますが、今のところ、トリアドバトラクスは、カエルとヒキガエルの系統樹の幹の近くに住んでいたことが知られている最古の先史時代の両生類です。この小さな生き物は、椎骨の数が現代のカエルとは異なり (現代の属の半分であるのに対し、14 個)、そのいくつかは短い尾を形成していました。しかし、そうでなければ、初期の三畳紀のトリアドバトラクスは、ぬるぬるした皮膚と強い後ろ足を備えた、明らかにカエルのような外形を呈していたでしょう。
32/33あちらへ

Nobu Tamura
今日まで、Vieraella の名声は化石記録の中で最古の真のカエルであるということです。ただし、長さ 1 インチ強、1 オンス未満の非常に小さなものではあります (古生物学者は、さらに古いカエルの祖先である「トリプル フロッグ」を特定しました)。トリアドバトラクスは、現代のカエルとは解剖学的に重要な点で異なっていた)。ジュラ紀初期にさかのぼるビエラエラは、大きな目を持つ古典的なカエルのような頭を持ち、その小さくて筋肉質の脚は印象的なジャンプを可能にしました。
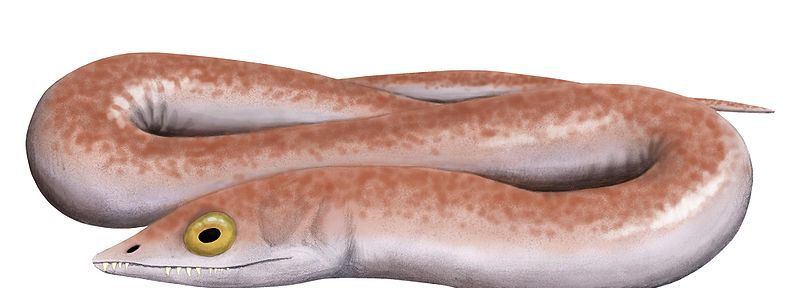
33/33ウェストロジアナ

Nobu Tamura
最も進化した先史時代の両生類が直接進化して最も進化していない両生類になったと言うのは、少し単純化しすぎています。 先史時代の爬虫類 ;硬い卵ではなく革のような卵を産む「羊膜類」として知られる中間グループもありました(したがって、水域に限定されませんでした)。初期の石炭紀のウェストロシアナは、古生物学者がその手首、椎骨、頭蓋骨の両生類のような構造に注目するまで、最古の真の爬虫類であると信じられていました(現在、ハイロノムスに与えられた名誉)。今日、ウェストロシアナがその後継者である真の爬虫類よりも原始的であったという無知な声明を除いて、この生き物をどのように分類するかについて完全に確信している人は誰もいません!